4. 干细胞与前体细胞
——高山流水
由于PRC1将谱系决定因子(lineage determination factor)基因组织在一起,维持它们待表达的状态,使基因调控网络(gene regulatory network)滞留在某个阶段,Hedgehog-Bmi1途径可以维持干细胞静态,使干细胞通过细胞质极性分裂持续产生前体细胞(progenitor),形成高山流水,即静态干细胞是上游源头,前体细胞和分化的功能细胞是下游。
一旦特定的谱系决定因子的表达被确立,它会将其下游的功能基因组织在一起。开始时,它受Rb和HDAC的抑制,当接受到Wnt分化信号后,会启动Myc的表达,Myc替换HDAC,功能基因表达,形成成熟的功能细胞。
谱系决定因子同样是染色质高级结构的组织者。与PRC2调控Hox基因簇和PRC1调控谱系决定因子基因不同,谱系决定因子调控的是行使具体功能的蓝领基因,并不属于基因调控网络。
Hox基因簇的多米诺式表达需要PRC2(催化组蛋白H3K27me3甲基化),MLL(催化组蛋白H3K4me3甲基化),RBP2(催化组蛋白H3K4me3去甲基化)。谱系决定因子基因的依次表达需要PRC1(催化组蛋白H2AK119ub1泛素化)。 功能基因的表达也需要组蛋白修饰酶的参与,只是另外一套:功能基因的持久抑制需要HP1(催化组蛋白H3K9me3甲基化);短暂抑制需要HDAC(催化组蛋白去乙酰化);表达需要SWI/SNF(染色质重塑复合物,即chromatin remodeler)。
PRC1将谱系决定因子基因组织在一起形成的结构,叫PcG (Polycomb Group) body。功能基因的抑制和激活也在细胞核的特定位置发生:抑制主要在核纤层关联域(LADs, lamin-associated domains);激活在转录工厂(transcription factory)。
干细胞的自我更新与分化
静态干细胞和前体细胞通过细胞质极性分裂产生两个细胞,一个继续维持原态,一个将要走向分化。 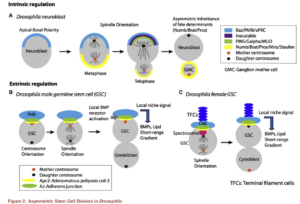 Asymmetric Stem Cell Division: Precision for Robustness –Cell stem cell 11: 461 (2012)
Asymmetric Stem Cell Division: Precision for Robustness –Cell stem cell 11: 461 (2012)
在染色体的极性分裂过程中,需要原来的和新合成的组蛋白(H3-H4)2,kinetochore,中心体的极性分布,它们在PRC2调控Hox基因簇的多米诺式表达中起重要作用。在静态干细胞分裂产生前体细胞,及前体细胞分裂产生功能细胞的过程中同样涉及极性分裂,但更多的是细胞质的极性分裂,需要aPKC,Numb等蛋白的极性分布,aPKC分布到未分化的细胞中,Numb分布到将分化的细胞中。
信号间的平衡
干细胞静态维持,自我更新,分化,取决于Shh,Growth Factor(GF,即EGF和FGF),Wnt,TGFβ,miRNA间的博弈。Shh激活Bmi1,维持干细胞静态,静态干细胞通过极性分裂,产生两个细胞,一个维持静态,另一个将要分化。在将要分化的细胞中,GF使它自我复制,Wnt激活Myc,Myc替换HDAC,使其分化,TGFβ促使细胞迁移,miRNA协调GF,Wnt和TGFβ间的平衡。
信号分子通过基因调控网络或转录因子使细胞产生特定的行为:shh通过Bmi1维持干细胞静态;Growth Factor通过E2F控制细胞分裂;Wnt通过Myc使前体细胞分化成功能细胞;TGFβ通过Snail或Twist控制EMT (Epithelial-Mesenchymal Transition)和迁移。
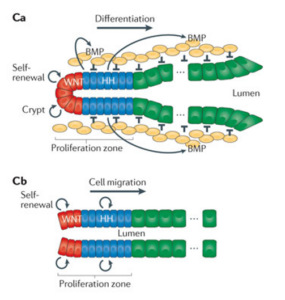 Mechanisms and functions of Hedgehog signalling across the metazoa –Nature Reviews Genetics 12: 393 (2011)
Mechanisms and functions of Hedgehog signalling across the metazoa –Nature Reviews Genetics 12: 393 (2011)
在肠上皮组织中,Transit-amplifying cells(即前体细胞)产生Shh信号分子,维持干细胞静态。
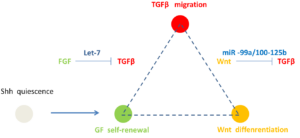 FGF regulates TGF-β signaling and endothelial-to-mesenchymal transition via control of let-7 miRNA expression –Cell reports 2: 1684 (2012)
FGF regulates TGF-β signaling and endothelial-to-mesenchymal transition via control of let-7 miRNA expression –Cell reports 2: 1684 (2012)
miR-99a/100-125b transcription regulate hematopoietic stem and progenitor cell homeostasis by shifting the balance between TGFβ and wnt signaling –Genes & Development 28: 858 (2014)
Let-7调节FGF和TGFβ间的平衡,miR-99a/100-125b调控Wnt和TGFβ间的平衡。
转录工厂
当干细胞分化成功能细胞后,PRC2调控Hox基因簇,非基因簇同源域蛋白的表达,及PRC1调控谱系决定因子逐渐退出舞台,除最终被选定表达的谱系决定因子基因外,其它与发育相关的转录因子基因被DNA甲基化抑制。 The polycomb group protein Ezh2 directly controls DNA methylation –Nature 439: 871 (2006)
谱系决定因子直接决定细胞的分化命运,它将下游的功能基因组装在一起,形成转录工厂(transcription factory)。这些功能基因编码行使具体功能的蓝领蛋白,而基因调控网络是指转录因子间的相互激活和抑制,所以谱系决定因子介导形成的染色质高级结构不属于基因调控网络,而是基因调控网络输出的最终结果。
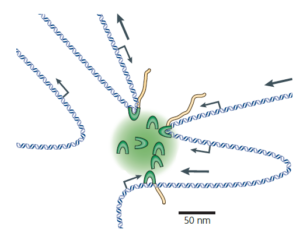 Transcription factories: gene expression in unions? –Nature reviews genetics 10: 457 (2009)
Transcription factories: gene expression in unions? –Nature reviews genetics 10: 457 (2009)
Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cell –Nature genetics 42: 53 (2010)
功能相关的基因被谱系决定因子组织在一起,由RNA聚合酶II统一转录,形成转录工厂。
核纤层关联域
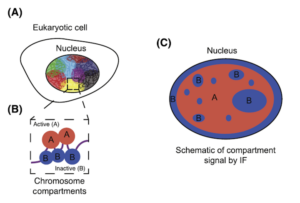 Schematic of subnuclear localization of A (active) and B (inactive) compartments. B compartments (blue) are spatially localized to the nuclear periphery and surrounding the nucleoli, while A compartments (red) are more interior.
Schematic of subnuclear localization of A (active) and B (inactive) compartments. B compartments (blue) are spatially localized to the nuclear periphery and surrounding the nucleoli, while A compartments (red) are more interior.
 Mechanisms and functions of chromosome compartmentalization –Trends in biochemical sciences 45: 385 (2020)
Mechanisms and functions of chromosome compartmentalization –Trends in biochemical sciences 45: 385 (2020)
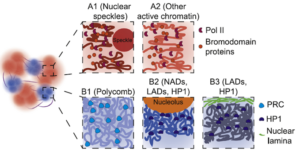
细胞核内,处于表达状态的基因位于内部,而不表达的基因围绕着核膜或核仁排列,即核纤层关联域(LADs, lamin-associated domains)或核仁关联域(NADs, nucleolar-associated domains)。
基因表达的区域富含RNA聚合酶II (pol II)和Bromodomain proteins (Bromodomain蛋白是染色质重塑复合物SWI/SNF的成员),形成转录工厂,主要负责功能基因的表达。位于LADs和NADs的基因含HP1(heterochromatin protein 1,可结合H3K9位点甲基化的组蛋白),主要是未表达的功能基因。受polycomb调控的基因位于细胞核内部,主要是受PRC2调控Hox基因簇,及受PRC1调控的谱系决定因子(lineage determination factor)基因。
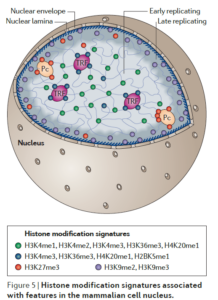
Charting histone modifications and the functional organization of mammalian genomes –Nature reviews genetics 12: 7 (2011)
H3K4位点的甲基化修饰集中在转录工厂(TRF,transcription factory),H3K27位点的甲基化修饰集中在Pc(PcG body),H3K9位点的甲基化修饰集中在核纤层关联域(LADs, lamin-associated domains)。
RB,HDAC,Myc
在核纤层关联域(LADs, lamin-associated domains)中,RB和HDAC会参与功能基因的抑制,但转录因子可以使基因离开抑制区而表达,所以RB和HDAC有点像PcG body中的Bmi1,是暂时抑制基因表达的,只是RB和HDAC调控的是功能基因,Bmi1调控的是谱系决定因子基因。
谱系决定因子的转录功能受RB和HDAC的调控,HDAC催化组蛋白去乙酰化修饰,抑制基因转录,Myc替换HDAC后,启动基因表达。
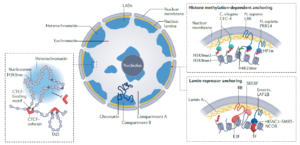 Establishment of H3K9-methylated heterochromatin and its functions in tissue differentiation and maintenance –Nature reviews molecular cell biology 23: 623 (2022)
Establishment of H3K9-methylated heterochromatin and its functions in tissue differentiation and maintenance –Nature reviews molecular cell biology 23: 623 (2022)
核纤层关联域通过两种机制抑制功能基因,一种是通过HP1(heterochromatin protein 1)结合H3K9me2,H3K9me3修饰的组蛋白,可以较稳定的抑制功能基因;另一种是通过RB和HDAC将功能基因暂时锚定在核纤层关联域,转录因子可以将基因激活,类似PcG body中,Bmi1将谱系决定因子基因暂时链接在一起。
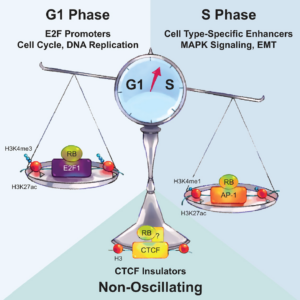
Chromatin-bound RB targets promoters, enhancers, and CTCF-bound loci and is redistributed by cell-cycle progression –Molecular Cell 82: 3333 (2022)
RB不仅能通过E2F1调控细胞分裂,还能参与谱系决定因子调控细胞分化的过程。
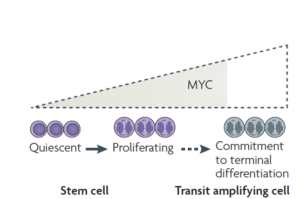 MYC in mammalian epidermis: how can an oncogene stimulate differentiation? –Nature reviews Cancer 8:234 (2008)
MYC in mammalian epidermis: how can an oncogene stimulate differentiation? –Nature reviews Cancer 8:234 (2008)
The opposing transcriptional functions of sin3a and c-myc are required to maintain tissue homeostasis –Nature cell biology 13: 1395 (2011)
c-myc regulates transcriptional pause release –Cell 141: 432 (2010)
Class I histone deacetylases sequentially interact with MyoD and pRb during skeletal myogenesis (MyoD是谱系决定因子,使细胞分化成肌肉细胞。) –Molecular cell 8: 885 (2001)
HDAC抑制谱系决定因子的转录功能。在干细胞分化成功能细胞的过程中,Myc替换掉HDAC,谱系决定因子启动功能基因的表达。Myc是Wnt信号的下游,所以Wnt信号主要促进细胞走向分化。
三类启动子
三类启动子的基因包括:一是发育转录因子基因(Developmental transcription factors),主要是受PRC2调控的Hox基因簇,非基因簇的同源域基因,及受PRC1调控的谱系决定因子基因;二是使细胞走向最终分化的功能基因(terminally differentiated cell-specific genes),就是谱系决定因子激活的蓝领蛋白基因;三是持家基因(housekeeping genes),就是类似actin的组成型表达的基因。
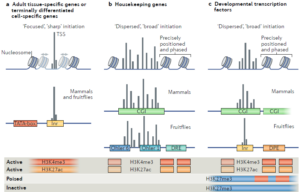 Eukaryotic core promoters and the functional basis of transcription initiation –Nature reviews molecular cell biology 19: 621 (2018)
Eukaryotic core promoters and the functional basis of transcription initiation –Nature reviews molecular cell biology 19: 621 (2018)
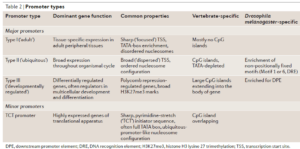 CpG island promoters is SWI/SNF-independent, whereas most promoters with SWI/SNF-dependent activation lack CpG islands.(SWI/SNF是染色质重塑复合物,即chromatin remodeler) Metazoan promoters: emerging characteristics and insights into transcriptional regulation –Nature reviews genetics 13: 233 (2012)
CpG island promoters is SWI/SNF-independent, whereas most promoters with SWI/SNF-dependent activation lack CpG islands.(SWI/SNF是染色质重塑复合物,即chromatin remodeler) Metazoan promoters: emerging characteristics and insights into transcriptional regulation –Nature reviews genetics 13: 233 (2012)
功能基因的启动子,不含GpC islands,需要SWI/SNF来协助表达;受PRC2和PRC1调控的发育转录因子基因的启动子,含GpC islands,不需要SWI/SNF协助表达。