1. 总结
——倚天照海花无数,流水高山心自知
The universe can be finite, but without walls, because if I go in one direction, I keep going, and I come back from the other direction.
宇宙可以是有限的,但没有墙,因为如果我朝一个方向前进,不断地前进,就会从另一个方向回来。–爱因斯坦 (纪录片《无限旅途》)
I could be bounded in a nutshell and count myself a king of infinite space.
即便被关在果壳之中,我仍然自以为是无限空间之王。–莎士比亚 《哈姆雷特》
一切美好的事物,都是在曲折中接近自己的目标。一切笔直都是骗人的,所有的真理都是弯曲的。时间本身即是一个圆圈。–尼采
对事物的认识,往往归结于对形状的理解。画家保罗.塞尚说,“世界上有各种不同的造型,可是任何的造型,最后还原到本质,其实就是数学几何学上的所谓的圆锥体,圆柱体,三角形,长方形”。画家葛饰北斋说,“这个世界是由圆圈和线条组成的–电影《眩 北斋之女》”。对生物的认识,也会归结于对染色质高级结构的理解。
三种染色质高级结构
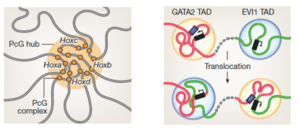 Transcription factors and 3D genome conformation in cell-fate decisions –Nature 569: 345 (2019)
Transcription factors and 3D genome conformation in cell-fate decisions –Nature 569: 345 (2019)
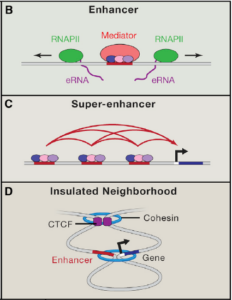 Transcriptional addiction in cancer –Cell 168: 629 (2017)
Transcriptional addiction in cancer –Cell 168: 629 (2017)
胚胎发育过程像一个历史舞台,不同的角色在不同的时间,地点依次登场。 原肠运动是同源域蛋白(homeodomain protein)起作用的阶段,间充质细胞中Hox基因簇由PRC2,MLL和RBP2 调控多米诺式表达,Hox基因通过信号分子决定与之结合的上皮细胞中非基因簇同源域蛋白(unclustered homeodomain protein)的表达,启动上皮细胞中基因调控网络(gene regulatory network)的特定分支(subcircuit),产生特定的干细胞。
干细胞中PRC1(Bmi1)通过染色质拓扑结构域(TAD,topologically associated domains)将谱系决定因子(lineage determination factor)基因组织成高级结构,调控它们依次表达。
基因调控网络中,多个转录因子会通过超级增强子(super-enhancer)的方式协同调控下游转录因子的表达,CTCF会参与超级增强子的功能。
就像《杯酒人生》中打开一瓶葡萄酒:我喜欢想酒的一生,把它想成是有生命的东西,想葡萄成长的那年发生了什么事,下过雨之后的太阳如何闪耀,想所有采集葡萄的人们,如果那是瓶老酒,他们又有多少人现已不在人世,要是我今天开了一瓶酒,它的味道会跟我改天开不一样,因为一瓶酒其实有生命,它会持续演变,变繁复,直到它达到最完美的境界,然后它会维持稳定,最后不可避免的衰老。 (I like to think about the life of wine, how it’s a living thing. I like to think about what was going on the year the grapes were growing, how the sun was shining, if it rained. I like to think about all the people who tended and picked the grapes, and if it’s an old wine, how many of them must be dead by now. I like how wine continues to evolve, like if I opened a bottle of wine today, it would taste different than if I’d opened it on any other day. Because a bottle of wines actually alive, and it’s constantly evolving and gaining complexity, until it peaks, and then it begins its steady inevitable decline.)
植物中没有Hox基因簇,也没有明确的PRC1基因。植物发育过程中有Wus-CLV1-STM途径,类似于动物中间充质Hox基因-信号-上皮组织中非基因簇同源域蛋白途径的mini版。植物中的Wus-CLV1-STM途径更多受小分子激素的调控,在发育过程中可被重复启动。
PRC2,PRC1和转录因子是染色质高级结构的三种组织者。在细胞分裂时,DNA需复制,染色质高级结构会被打开,染色质高级结构组织者的一个重要特性是:通过染色质上的书签(bookmark),在细胞分裂后能重新构建染色质高级结构。PRC2和PRC1使用的书签是经过修饰的组蛋白,而转录因子本身就是书签,有些转录因子在细胞分裂时,会一直结合在染色质上。 Mitotic bookmarking by transcription factors –Epigenetics & Chromatin 6: 6 (2013)
动物和植物的基因调控网络的等级性
基因最初是孟德尔做豌豆的遗传实验时提出的概念,孟德尔通过观察豌豆某个性状的改变在子代的分布比例,推算出应该存在可以在父代和子代间传递的遗传物质,即基因,基因突变决定了性状的改变,但当时并不知道遗传物质是DNA。 (注:是基因突变决定性状改变,不是基因决定性状。性状是发育来的,是蛋白和细胞协同作用的结果,不是单个基因能决定的。基因突变决定性状改变,也需依赖特定的遗传背景才能成立。)
进一步观察两个性状的改变在子代间的分布比例,发现有些基因是连锁在一起的,有些基因是相互独立的,即产生了染色体的概念。
一个基因突变决定某个性状改变,不会影响另一个性状的改变,如果两个性状的改变在子代的出现总是绑定的,就可以推理出两个基因是连锁的,即系统本身的重点性决定了整体改变内因的独立性,也就是封面上那四个圆想表达的意思。
分子遗传学,细胞生物学,生物化学把单个细胞讲得很清楚了,但细胞如何组建成多细胞个体,即发育学是生物中最难理解的部分。
发育离不开三个基本概念:基因调控网络的等级性(原动力);间充质与上皮细胞(板块的相互作用);成体干细胞(山的形成与高山流水)。
基因调控网络的等级性是发育的原动力,最初的胚胎干细胞位于最上端。
大部分器官(大脑,心脏等除外)是由间充质细胞和上皮细胞共同构建的,这种相互作用是在原肠运动中发生的。原肠运动时,间充质细胞中的PRC2调控Hox基因簇的多米诺式表达,为发育提供了固定的框架,使与之结合的上皮细胞走向基因调控网络的不同分支,并产生成体干细胞。
成体干细胞是由PRC1将谱系决定因子基因链在一起,使基因调控网络滞留在某个阶段,并由干细胞微环境来维持,使之可以持续地依次产生功能细胞。
高等动物的基因调控网络的等级性是不可逆的,失去再生能力,而植物的基因调控网络更多受小分子激素控制,是可逆的,所以才会“野火烧不尽,春风吹又生–唐代诗人白居易”。
植物没有原肠运动,没有Hox基因簇,也就没有发育中的固定框架。植物的发育受外界环境影响,光照,温度都会决定植物是否分化出花。
植物没有严格意义上的PRC1,也没有严格意义上的干细胞。植物的部分细胞具有全能性,如水稻的下胚轴或一些观赏植物的叶子,都可以通过组培中的激素比例调整,长出愈伤,并发育成一株完整的个体。
单细胞的微生物只有细胞分裂,没有细胞分化,多细胞植物和动物有细胞分化,也就有了受基因调控网络控制的发育过程。生物的进化过程和地球的演变过程相对应,地球最初表面只有水,产生了单细胞的微生物,然后有了风,水面上出现了海浪,进化出了植物,后来出现了大陆,板块运动产生了山,生物也进化出了Hox基因簇和PRC1,出现了动物。
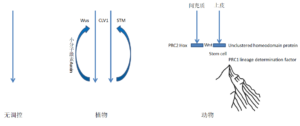 在没有任何调控机制的情况下,基因调控网络的等级性类似于一个铁球垂直落向地面。
在没有任何调控机制的情况下,基因调控网络的等级性类似于一个铁球垂直落向地面。
植物中没有Hox基因簇,没有PRC1,基因调控网络具有可逆性,小分子激素,如生长素(Auxin)可以使其重复的返回到上游,所以植物的发育过程类似于一波波的海浪。
动物中两个组织的相互作用,形成了干细胞微环境,间充质细胞转变成成纤维细胞或基质细胞,维持Hox基因表达模式,不再走向下游,在与之相结合的组织中非基因簇的同源域蛋白决定干细胞的分化属性,干细胞中,由PRC1将谱系决定因子基因组织在一起,维持它们待表达的状态,使基因调控网络滞留在某个阶段,分化过程中调控它们依次表达。如果把PRC2调控Hox基因簇多米诺式表达比喻成地球板块的形成,非基因簇的同源域蛋白起作用的阶段就是板块间的相互作用,干细胞中PRC1将谱系决定因子基因组织成染色质高级结构就是造山运动,干细胞自我更新及依次分化产生功能细胞的过程就是高山流水。 植物和动物基因调控网络的区别就像晚清重臣曾国藩的一幅对联:倚天照海花无数,流水高山心自知(前句来源于宋代诗人苏轼,后句来源于宋代诗人王安石)。