3. PRC1与谱系决定因子
——山的形成
间充质细胞中Hox基因簇的多米诺式表达,为形态建成提供了固定框架,决定了干细胞出现的位置。干细胞中特定的非基因簇同源域基因(unclustered homeodomain gene)的表达,启动基因调控网络(gene regulatory network)特定的分支(subcircuit),决定了干细胞的分化方向。
干细胞中,非基因簇同源域蛋白将待表达的谱系决定因子基因(lineage determination factor)组建成的PcG body,canonical PRC1成员 Bmi1将基因链接起来,位于边缘的谱系决定因子基因优先表达。不表达的谱系决定因子基因由yin yang 1招募non-canonical PRC1而被稳定抑制。
PRC1(Bmi1)维持谱系决定因子基因待表达的状态,使基因调控网络的等级性(hierarchy)滞留在某个阶段,是干细胞存在的内在因素。Shh信号可以激活Bmi1的表达,使干细胞保持静态,是干细胞存在的外在条件。

《创世纪》米开朗基罗
人是上帝创造出来最美的艺术品。 –柏拉图
PRC2和PRC1像神的两只手,塑造了高级的生命。
PRC2调控的是Hox基因簇;而PRC1通过染色质高级结构将谱系决定因子基因组织在一起,它们在染色体上可以距离很远,也可以不在一条染色体上。 多米诺的形成是一次性的,发生在胚胎发育初期,如原肠运动;因为有静态干细胞的维持机制,PRC1调控谱系决定因子基因依次表达,是持续的。 PRC2调控Hox基因簇的多米诺式表达是与DNA复制及染色质极性分裂相偶联的,需PRC2,MLL和RBP2的协同作用来实现;而PRC1调控谱系决定因子基因的依次表达,是通过接受外界信号和PRC1的成员替换来实现的,PRC2并不是必须的。 PRC2调控Hox基因簇的区域化表达是确定框架,是楼房的结构;PRC1调控谱系决定因子的依次表达是形成功能细胞,是内部装修。如果前者是毛毛虫,后者则是蝴蝶。
染色质拓扑结构域
基因调控网络(gene regulatory network)的等级性(hierarchy),一种是由转录因子间相互激活和抑制形成的逻辑关系决定的,另一种是由染色质的高级结构形成的,其中包括PRC2调控Hox基因簇的多米诺式表达,和PRC1(Bmi1)将谱系决定因子(lineage determination factor)基因通过染色质拓扑结构域(TAD,topologically associated domains)的形式构建在一起。
如果只有第一层面的逻辑关系形成的等级性,那么就像是电脑中由二级管,三级管组成的与,或,非门,生命体的形成变成了按照事先编程好的程序运行的结果,其实不然,因为有染色质高级结构的参与,使生命体的发育更像是一座雕塑,一副画的创作,使它成为一件艺术品。就像爱因斯坦所说,“逻辑会把你从A带到B,想象力能带你去任何地方”。
关于艾伦.图灵(Alan Turing)的纪录片《密码破译者》中图灵的一段独白,“大多数花的花瓣数都是斐波那契数,每一个数都是前两位数之和,1 1 2 3 5 8 13 21 34,细胞自身没有大脑,但细胞群却会分裂,最终,一些细胞变成鸟的脊椎骨,其他一些细胞则变成了翅膀,那么单细胞是如何知道它要变成什么器官呢?因此我问自己,自然界的存在模式是否具有数学理论基础,比如牛身上的斑点,或这朵雏菊的花瓣。数学家们会谈起数学的美丽,但他们谈的是可计算数,是一种解决问题的美,那些更大的无穷,那些不可计算物,就在人类虚弱的意识所征服的微小知识外。” (For some mad reason, the number of petals on most flowers is a Fibonacci number. Each number the sum of the previous two: 1, 1, 2, 3, 5, 8, 13, 21, 34. Cells don’t have a mind of their own, but a clump of cells will split, and eventually some of the cells will become the backbone of a bird, and some of the others will become its wings. But how do the individual cells know what part of the organism to become? So I asked myself whether there might be some mathematic underpinning to patterns that occur in nature, like the spots on a cow, or the petals on this daisy. Mathematicians talk about the beauty of numbers, they’re talking about the computable, the beauty of what can be resolved. But what about the rest, the greater infinity? What’s not computable lies beyond the infinitesimal sliver of knowledge. We’ve managed to subdue inside our fragile, trembling human consciousness.)
如果说基因调控网络的等级性是生物形态建成的基础,那么转录因子相互激活和抑制形成的逻辑关系属于可计算部分,而染色质高级结构层面的调节就是不可计算的部分。
 Organization and function of the 3D genome –Nature reviews genetics 17: 661 (2016)
Organization and function of the 3D genome –Nature reviews genetics 17: 661 (2016)
The role of 3D genome organization in development and cell differentiation –Nature reviews molecular cell biology 20: 535 (2019)
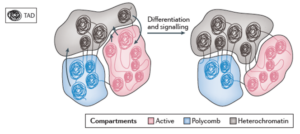
TAD是通过酶切和测序等技术手段能够检测到的染色质loop结构,染色质在TAD的基础上可以进一步被组织成激活区(Active),待激活区(Polycomb),和抑制区(Heterochromatin),PRC1是Polycomb bodies的组织者,在Polycomb bodies中基因处于待表达的状态。
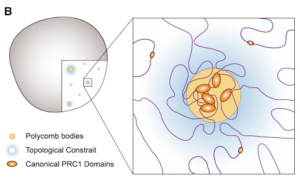 We propose that PRC1-bound loci cluster together in the nucleus to form discrete polycomb bodies. (PRC1是polycomb bodies的主要组织者。)
We propose that PRC1-bound loci cluster together in the nucleus to form discrete polycomb bodies. (PRC1是polycomb bodies的主要组织者。)
A central role for canonical PRC1 in shaping the 3D nuclear landscape –Genes & development 34: 931 (2020)
PRC2 does not seem to be critical for establishing TADs, and its depletion has only limited effects on long-range interactions of Polycomb domains. In contrast, PRC1 plays a critical role, both in local chromatin condensation of Polycomb domains and in the establishment of their long-range interactions. (PRC1将谱系决定因子基因组织在一起的过程中,PRC2不是必须的。)
Genome regulation by polycomb and trithorax: 70 years and counting –Cell 171: 34 (2017)
由于PRC2对于TAD的形成不是必须的,所以PRC1调控谱系决定因子的依次表达,不可能通过PRC2调控HOX基因簇多米诺式表达的那种与细胞分裂相偶联的自主机制,只能通过接受外界信号来完成。
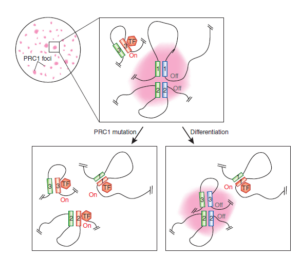 PRC1 proteins orchestrate three-dimensional genome architecture –Nature genetics 47: 1105 (2015)
PRC1 proteins orchestrate three-dimensional genome architecture –Nature genetics 47: 1105 (2015)
Polycomb repressive complex 1 generates discrete compacted domains that change during differentiation –Molular cell 65: 432 (2017)
在干细胞依次分化的过程中,由于染色质的高级结构,特定的染色质拓扑结构域基因被依次激活。
转录因子与PcG body
Coherent Hox complexes are expressed in a vectorial or sequential fashion with respect to the coordinates of body.
They act as swithes that allow GRN (gene regulatory network) patterning subcircuits in given locations of the body plan, or alternately they prohibit these subcircuits in given locations.
Evolution of gene regulatory networks controlling body plan development –Cell 144: 970 (2011)
同源域蛋白在基因调控网络的等级中起承上启下的作用,一方面Hox基因簇的多米诺式表达提供整体框架,另一方面非基因簇同源域蛋白协助canonical PRC1 (Bmi1)将特定的谱系决定因子基因组装成PcG body,从而启动基因调控网络的特定分支。
We propose that cell fate transcription factors are strong candidates for the regulation of PcG protein recruitment to and dissociation from their target genes.
Polycomb group proteins: navigators of lineage pathways led astray in cancer –Nature reviews cancer 9: 773 (2009)
非基因簇同源域蛋白(及pioneer transcription factor)参与PcG (Polycomb Group) body的组建,PcG body是canonical PRC1将谱系决定因子基因组织在一起的结构。
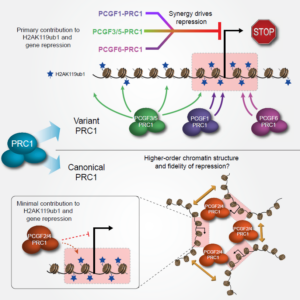
Synergy between Variant PRC1 Complexes Defines Polycomb-Mediated Gene Repression –Molecular cell 74: 1020 (2019)
Chromatin folding and nuclear architecture: PRC1 function in 3D –Current Opinion in Genetics & Development 55: 82 (2019)
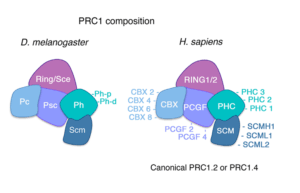
非基因簇同源域蛋白特异地选择某些谱系决定因子基因,构建PcG body,其中含Bmi1,也就是canonical PRC1。其他的谱系决定因子基因由普遍存在的转录因子YY1 (yin yang 1)招募non-canonical PRC1来抑制,含RYBP (RING1-Yin Yang1 (YY1) Binding Protein),不组装成PcG body。
PRC1与Bmi1
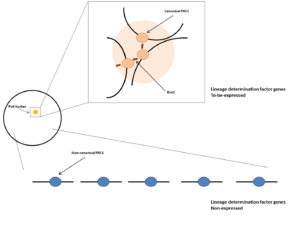 特定的unclustered homeodomain蛋白或pioneer transcription fator激活基因调控网络(gene regulatory network)某一个分支,使某些谱系决定因子基因被canonical PRC1(主要由Bmi1,即PCGF4链接)组织在一起,形成PcG body结构,位于边缘的谱系决定因子基因率先解开而表达。而其它分支的谱系决定因子基因被Non-canonical PRC1抑制。
特定的unclustered homeodomain蛋白或pioneer transcription fator激活基因调控网络(gene regulatory network)某一个分支,使某些谱系决定因子基因被canonical PRC1(主要由Bmi1,即PCGF4链接)组织在一起,形成PcG body结构,位于边缘的谱系决定因子基因率先解开而表达。而其它分支的谱系决定因子基因被Non-canonical PRC1抑制。
如果把PcG body的形成比喻成山的形成,那么在PcG body中的待表达的谱系决定因子基因就是位于山上的,越先表达的基因海拔越高,而被YY1普遍抑制的谱系决定因子基因就是海平面。
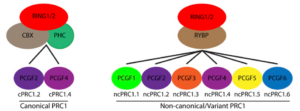 The genetic basis for PRC1 complex diversity emerged early in animal evolution –PNAS 117: 22880 (2020)
The genetic basis for PRC1 complex diversity emerged early in animal evolution –PNAS 117: 22880 (2020)
Canonical PRC1包括Bmi1(PCGF4),CBX和PHC。Non-canonical PRC1包括PCGF1-6和RYBP (RING1-Yin Yang1 (YY1) Binding Protein)。
基因组印记,Hedgehog-Bmi1途径
基因组印记(Genomic imprinting)是指来自父方和母方的等位基因具有不同的表达特性。基因组印记只是PRC1(Bmi1)介导谱系决定因子基因形成染色质高级结构的副产物,很多与发育相关的基因都有基因组印记现象。 Genomic imprinting and physiological processes in mammals –Cell 176: 952 (2019)
受基因组印记调控的基因和谱系决定因子基因一样,在静态干细胞中,由PRC1(Bmi1)组装成染色质高级结构,当来自父母本的等位基因序列不同,被组装的高级结构不同时,就表现为基因组印记现象。Bmi1和基因组印记都可以作为静态干细胞的标志。
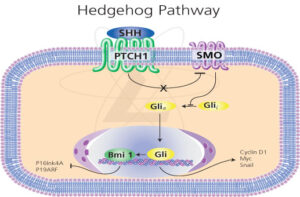 Hedgehog-Bmi1 pathway(from Sigma-aldrich)
Hedgehog-Bmi1 pathway(from Sigma-aldrich)
Lung stem cell self-renewal relies on Bmi1-dependent control of expression at imprinted loci –Cell stem cell 9: 272 (2011)
Hedgehog activity maintains adult lung quiescence and regulates repair and regeneration –Nature 526: 578 (2015)
Maternal imprinting at the h19-igf2 locus maintains adult haematopoietic stem cell quiescence –Nature 500: 345 (2013)
在肺上皮组织的静态干细胞,和静态的造血干细胞中存在Bmi1依赖的基因组印记。干细胞静态维持所需的外部信号是Shh,它通过Gli1激活Bmi1,Bmi1将谱系决定因子基因组织成染色质高级结构。Hedgehog-Bmi1 途径将干细胞静态,Bmi1,基因组印记三者联系起来。
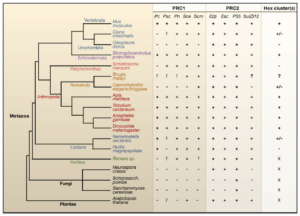 Genome regulation by polycomb and trithorax proteins –Cell 128: 735 (2007)
Genome regulation by polycomb and trithorax proteins –Cell 128: 735 (2007)
人(Mus)和果蝇(Drosophila)含有PRC1,PRC2和Hox基因簇。在线虫(Elegans)中只有部分的Hox基因簇。以拟南芥(Arabidopsis)为代表的植物中,有PRC2,但没有PRC1和Hox基因簇。
 Emerging roles for phage separation in plants –Developmental cell 55: 69 (2020)
Emerging roles for phage separation in plants –Developmental cell 55: 69 (2020)
植物没有PRC1,没有PRC1将谱系决定因子基因组织在一起的PcG body结构。
How Polycomb-mediated cell memory deals with a changing environment –BioEssays 40: 1700137 (2018)
(The Drosophila PRC1 core component can cause compaction of a nucleosome array in a Psc-dependent manner) Polycomb group proteins: repression in 3D –Trends in Genetics 27: 454 (2011)
Genomic imprinting absent in drosophia melanogaster adult females –Cell reports 2: 69 (2012)
(Imprinting has been detected in the endosperm and not the embryo) Cellular programming of plant gene imprinting –Cell 132: 735 (2008)
(Both C.elegans and A.thaliana do not encode a CTCF homolog) Ever-changing landscapes: transcriptional enhancers in development and evolution –Cell 167: 1170 (2016)
果蝇没有基因组印记,果蝇没有Bmi1基因,只有与之同源的Psc基因,Psc作为Canonical PRC1的成员,是构建PcG body的关键蛋白。植物中没有PRC1基因,胚发育中没有基因组印记机制。植物和线虫没有CTCF基因,CTCF蛋白参与染色质拓扑结构域的形成。
依次分化
sequential progenitor diversification model
Shaping our Minds: Stem and Progenitor Cell Diversity in the Mammalian Neocortex –Neuron 77: 19 (2013)
 Epigenetic control of adult stem cell function –Nature reviews molecular cell biology 17: 643 (2016)
Epigenetic control of adult stem cell function –Nature reviews molecular cell biology 17: 643 (2016)
Context-specific polycomb mechanisms in development –Nature reviews genetics 23: 680 (2022)
PRC1将谱系决定因子基因组织在一起的结构叫PcG body,是在显微镜下可见的细胞核内的结构。与PRC2将Hox基因簇组织在一起的微观结构相比,PRC1是在更大的空间将谱系决定因子基因组织在一起。推测PcG body类似一个湖泊,PRC1是船,每个PRC1携带一个谱系决定因子基因,而Bmi1通过锁链的机制将PRC1连在一起,这样位于边缘的谱系决定因子基因更容易脱离而表达,Bmi1也随之离开,第二个位于边缘的基因获得表达机会,以此类推形成依次表达。这使得Bmi1成为静态干细胞的标志蛋白,Bmi1的含量越高,细胞的分化能力越强。
 Mechanisms of bone development and repair –Nature reviews molecular cell biology 21: 696 (2020)
Mechanisms of bone development and repair –Nature reviews molecular cell biology 21: 696 (2020)
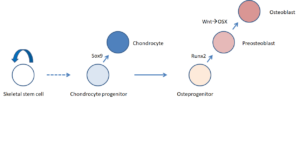
前体细胞依次分化出软骨细胞和成骨细胞的过程中,谱系决定因子基因Sox9和Runx2依次表达。
 Roadmap to embryo implantation: clues from mouse models –Nature reviews genetics 7: 185 (2006)
Roadmap to embryo implantation: clues from mouse models –Nature reviews genetics 7: 185 (2006)
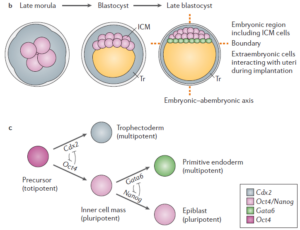
受精卵在最初的分化中表现为依次分化。
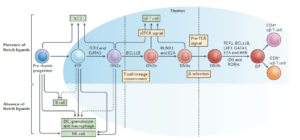 Developmental gene networks: a triathlon on the course to T cell identity –Nature reviews immunology 14: 529 (2014)
Developmental gene networks: a triathlon on the course to T cell identity –Nature reviews immunology 14: 529 (2014)
前体细胞先分化出B细胞,进入胸腺后再分化出T细胞。
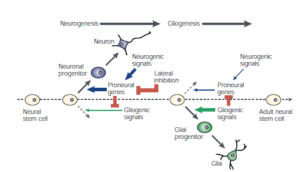 Generally, neural stem cells first generate neurons, and later produce glia, and the switch from neurogenesis to gliogenesis is the result of changes in stem-cell properties that are controlled by both extrinsic and intrinsic cues.
Generally, neural stem cells first generate neurons, and later produce glia, and the switch from neurogenesis to gliogenesis is the result of changes in stem-cell properties that are controlled by both extrinsic and intrinsic cues.
Proneural genes and the specification of neural cell types –Nature reviews neuroscience 3: 517 (2002)
神经细胞的前体细胞先分化出神经原,再分化出神经胶质细胞。
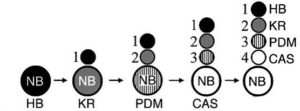 Drosophila neuroblasts sequentially express transcription factors which specify the temporal identity of their neuronal progeny –Cell 106: 511 (2001)
Drosophila neuroblasts sequentially express transcription factors which specify the temporal identity of their neuronal progeny –Cell 106: 511 (2001)
Neuronal subtype specification within a lineage by opposing temporal feed-forward loops –Cell 139: 969 (2009)
参禅之初,看山是山,看水是水
禅有悟时,看山不是山,看水不是水
禅中彻悟,看山仍然是山,看水仍然是水
–青原行思
人们习惯用已有的知识解释新的事物,而结果是已有的知识跟新的现象相关,但又不能准确解释,新的现象背后的机制是另一个层面上的,需再上一个台阶才能理解的东西。最开始想用多米诺机制来解释果蝇神经细胞转录因子的依次表达,后来觉得应该用PRC1调控谱系决定因子表达的机制来解释。多米诺调控的是Hox基因簇的依次表达,为胚胎发育提供位置信息,而PRC1 控制的是谱系决定因子,是在最终发挥功能的上皮细胞,神经细胞,造血细胞中起作用的。